
![]()
“Why is it so difficult for virologists to simply explain basic questions about a ‘virus’ such as whether the ‘virus’ is living or dead? Why must the concept of what a ‘virus’ is change depending upon the researchers and technology of the time? What physical organism changes in concept after over a century of supposed study? The answer to all of these questions is actually fairly easy to grasp. As the researchers have never actually had any physical entities on hand in order to study, the concept of what the assumed invisible entities are was allowed to constantly change in order to suit the needs and evidence of the researchers of the time. There was no solid foundation for virology to stand upon from the very beginning in order to definitively state what the nature of a ‘virus’ truly is.”
![]()
Inventing the Nature of “Viruses”
by Mike Stone, Viroliegy
November 10, 2023
“No viruses have been found multiplying free in nature.”
-Virologist Thomas Rivers
Tom Rivers: reflections on a life in medicine and science : an oral history memoir
For the greater part of the first 50 years of the 20th century, there was no agreed upon definition for what the invisible entities labelled as a “virus” actually were nor how these agents looked, formed and functioned. Some researchers believed that these entities were endogenous processes produced within the host while others envisioned them as exogenous invaders that came from outside and attacked from within. There were arguments over whether “viruses” were corpuscular in nature or whether they were a soluble liquid. Debates centered around whether these agents were alive or if they were simply inanimate and non-living. While there were researchers who believed “viruses” were a ferment or a chemical molecule of some kind, the majority believed that these invisible entities were just smaller unseen bacterium. According to biochemist and historian of science Ton van Helvoort’s 1996 paper When Did Virology Start?, the “virus” concept lacked clarity and certainty over the first half of the 20th century. However, the link between bacteriology and “viruses” was so strong at this time that these unseen entities were not considered conceptually distinct from bacteria:
“I have come to believe that, despite its widespread appearance in textbooks and journals of that era, the early concept of the “filterable virus” lacked clarity and certainty. More importantly, I also believe that during the 1930s and 194Os, the links between the study of filterable viruses and bacteriology were so strong that viruses were still considered merely another form of bacteria-not conceptually distinct, as they now are.”
The reason for these many contradictory ideas about the nature of the “virus” was a direct result of the fact that the researchers never had a physical entity on hand in order to study. The “virus” was nothing more than a fluid concept that was open to the interpretation of those who claimed to be working with them. Most of these researchers came from a bacteriological or chemistry background, and thus, they viewed the “virus” concept through their own lens and paradigms. Regardless, there was no way to actually determine the true nature of something that could not be seen or studied in reality and that only existed within the realm of the imagination.
Thus, it shouldn’t be hard to understand why virologists often have a difficult time answering simple questions such as “What is a virus?” or “Is it alive or dead?” This is exactly the argument made in the appropriately titled 2014 article Inventing Viruses by William Summers, a retired Professor of Therapeutic Radiology, Molecular Biophysics & Biochemistry, and History of Medicine. While being able to define what a “virus” is should be an easy task for any virologist, simple questions about the nature of a “virus” are not ones that are simple for them to answer. In the opening of his paper, Summers asked a more subtle question about the invention of the “virus” category:
“…how generations of microbiologists arrived at the idea that some of the entities they dealt with fell into a category that differed in fundamental ways from others. In other words, how did they invent the category of “virus” as we now know it?”
Summers looked to investigate how the idea that “viruses” are a separate entity that requires its own category away from bacteriology came to be. In doing so, he admitted that our beliefs, understandings, and conceptions of what a “virus” is changes over time. This is because “viruses” are whatever a virologist tells us that they are. The concept and the nature of the “virus” was invented, and continually reinvented, by virologists as part of the normal progress of their (pseudo) science. In other words, the idea of the “virus” is able to change at any time based upon whatever a virologist wants a “virus” to be at any given moment:
“Even so, how did the category “virus” come to be recognized, and what are its essential, defining qualities? Viruses are natural objects, but our beliefs, understanding, and conceptions of them change over time on the basis of new information, new points of view, and new scientific values and standards. In a very real way, a virus is what virologists say it is. It is a product of the way virologists talk about viruses—that is, the way facts about viruses are organized in their discourse. It can be said that virologists invent (and continually reinvent) the concept of a virus as part of the normal progress of their science.”
The deliberate ever-changing concept of the “virus” shifted away from its original invention as an agent of disease transmission to its modern day concept as a genetic assembly that sometimes causes disease when it integrates into its host in order to survive. This reinvention of the concept happened in 1957 when French microbiologist Andre Lwoff took many competing and contradictory ideas and mashed them together into the modern definition of a “virus” based upon work done with bacteriophages. Prior to his reinvention of the concept, in 1953, Lwoff actually questioned whether a bacteriophage was a “virus” and wanted to know exactly what a “virus” was. He even noted that “viruses” are defined to be exogenous (coming from outside of the body) while bacteriophages are “always formed inside its host” and “could therefore be described as endogenous,” i.e. originating from within the host. In fact, Lwoff stated that “if prophage is phylogenetically endogenous, the temperate phage produced by a lysogenic bacterium must be described as endogenous,” meaning that the phage is from within the host, thus negating it as an exogenous entity in line with the definition of a “virus.” Ironically, after redefining the “virus” as a genetic code in 1957, Lwoff would ultimately warn in 1991 that virology was “in danger of losing its soul, since viruses now show a strong tendency to become sequences.” He also argued that the abundance of discoveries was causing “the very concept of virus” to waver “on its foundations,” noting that the “problem today and in future is to keep abreast of its whereabouts.”
Regardless, Summers stated that his paper was not about the “triumphant accumulation of knowledge by the heroic scientists” of the past. Rather, it was an examination of the “continual struggle to understand and organize observations.” This struggle was showcased by Lwoff’s own attempts to rationalize and combine contradictory evidence in order to create the modern genetic concept of the “virus” from an entity that did not meet the necessary requirements:
“Nobelist Andre Lwoff, perhaps in a Gertrude Stein frame of mind, famously answered “viruses are viruses” (9), but the question “What is virus?” has been notoriously fraught since the role of virus in late nineteenth-century germ theories became central to medicine, and later, in the midtwentieth-century, to biology in general. The evolution, or perhaps deliberate and continuous reformulation, of the meaning of “virus” from an agent of disease transmission in the nineteenth century to a molecular assembly with remarkable properties by the end of the twentieth century is the subject of this article. This is not a story of the triumphant accumulation of knowledge by the heroic scientists of the past so much as it is an examination of the continual struggle to understand and organize observations that challenged and made obsolete the comfortable certainties of the often recent past. This examination requires consideration of past science on its own terms, without judgment in light of present-day understanding, and it requires consideration of the context and extent of background knowledge of the particular period considered.”
This struggle to answer the question “What is a virus?” was ongoing, even in the so-called “modern age” of virology. There was no consensus as to the true nature of a “virus.” Summers shared a quote by Joseph Beard that stated that the “virus” was a fabric of concepts that had been “woven of a plethora of woof and a paucity of warp.” In weaving terms, this makes for an unstable foundation upon which to weave. Another example was of plant virologist N.W. Pirie who was considered “agnostic” (impossible to know one way or the other) on whether a “virus” was a molecule or a microbe. However, he seemed to argue that the variability in the chemical composition of the same “virus” went against the modern molecular hypothesis. Thus, we can see that there was no agreement on the nature of the “virus:”
“The construction of the virus as a living molecule in the middle decades of the twentieth century generated wide debate as to the correct answer to the question, “What is a virus?” Having rejected filterability, negative growth properties, and size as defining characteristics, microbiologists searched for new ways to think about viruses. Even at the beginning of what might be called the modern era, there was remarkably little consensus on this subject. Joseph Beard, in 1945, famously remarked, “Viruses are said to be living molecules and autocatalytic enzymes and are likened to genes and mitochondria—in short, a fabric of concepts has been woven of a plethora of woof with a paucity of warp” (quoted in 47, p. 332). N.W. Pirie, one of the pioneers in the study of plant viruses, even in 1949 was agnostic as to whether viruses were microbes or molecules. In a long review of the problem in the British Medical Bulletin (47), he argued that the variation in chemical composition reported for the same virus suggested a level of heterogeneity not compatible with the molecular hypothesis. He noted that “all the viruses purified so far have contained nucleoprotein, but this generalization may lack significance because the viruses that have been studied are a group selected to some extent on a chemical basis.”
Summers ultimately concluded that each generation of virologists will look at “viruses” in their own way and will alter the concept of the “virus” based upon the “science” of the time. Thus, the “virus” is left to be a concept that is allowed to be continually reinvented at the whims of the researchers:
“Although “viruses are viruses,” each generation of scientists looks anew at these fascinating entities in its own way, endowing them with properties, relationships, and capacities that reflect the science of the time. Truly, they are microbes being continually reinvented by their most ardent admirers.”
In his summary, Summers laid out 5 very revealing points to end his paper on. Sharing similar sentiments as van Helvoort, he stated that the “virus” concept is an unstable one that “evolved,” not due to an accumulation of facts, but rather due to an ongoing reformulation of the “virus” concept on the basis of “scientific” focus at a given time. This reinvention was determined by technological advances rather than scientific understanding. Thus, the answer as to what a “virus” is will depend upon the discourse at the time more so than the “known” characteristics of “viruses:”
- The concept of a virus has not been stable and has evolved since its introduction in the latter half of the nineteenth century.
- This evolution has been not a linear accumulation of facts but rather an ongoing reformulation of the virus concept on the basis of scientific focus at a given time, e.g., growth, metabolism, chemical composition, genetics, or physical structure.
- The concept of a virus has particularly been determined by technological advances ratherthan scientific understanding.
- The answer to the question “What is a virus?” is one that depends on the particular scientific discourse at a given time.
- The discourse with respect to the physical object “virus” is based on the particular concerns and problems of interest at a given time more than on any one set of intrinsic characteristics known about viruses.
Why is it so difficult for virologists to simply explain basic questions about a “virus” such as whether the “virus” is living or dead? Why must the concept of what a “virus” is change depending upon the researchers and technology of the time? What physical organism changes in concept after over a century of supposed study? The answer to all of these questions is actually fairly easy to grasp. As the researchers have never actually had any physical entities on hand in order to study, the concept of what the assumed invisible entities are was allowed to constantly change in order to suit the needs and evidence of the researchers of the time. There was no solid foundation for virology to stand upon from the very beginning in order to definitively state what the nature of a “virus” truly is.
While Summers paper on the invention of the “virus” offers some great modern insight into the problems related to defining the nature of the invisible beast, there is a much earlier paper by prominent virologist Thomas Rivers from 1932 that details the many issues with trying to give life to the imaginary shortly after its conception. You may know Rivers due to his 1937 proclamation that “It is obvious that Koch’s postulates have not been satisfied in viral diseases.” This shockingly honest admittance that the essential logical criteria considered necessary in order to prove a microbe causes disease remains unfulfilled for “viruses” and continues to haunt virology to this day. As it is a rather long 18 pages that I have reproduced here, I will try to keep my commentary throughout brief. However, what Rivers highlighted as key problems in 1932 during the formative years of virology compliments Summers 2014 paper on why virologists needed to invent, and then continually reinvent, the concept of the “virus” that was dreamt up in the late 1800s.
Thomas Rivers immediately began his 1932 paper on the nature of “viruses” by admitting that, up to 1932, “viruses” were defined solely based upon their absence as well as for what they were not. “Viruses” were defined in negative terms as they were:
- Invisible to ordinary microscopic methods.
- Unable to be obtained via filtration.
- Unable to propagate in the absence of susceptible cells.
Interestingly, things did not progress away from defining “viruses” in negative terms even with Andre Lwoff’s 1957 modern reinvention of the concept as noted by Professor Milton W. Taylor, teacher of virology and world-renowned historian from Indiana University. In a 2014 paper examining what a “virus” is, Taylor explained that Lwoff’s reinvention of the “virus” concept was also a “negative definition” that “stresses the non-cellular nature of viruses.” By Lwoff’s own words from his 1971 paper From Protozoa to Bacteria and Viruses. Fifty Years with Microbes, he defined “viruses” by the “inability to grow and to divide, absence of metabolism, absence of the information for the enzymes of energy metabolism…the absence of transfer RNA and of ribosomes and also of the corresponding information.” In other words, even by the modern definition, “viruses” were still defined by what they were not.
While Rivers attempted to define “viruses” in what he felt were positive terms of what was “definitely known” about these invisible agents, he admitted that the biological nature was still a moot question, i.e. one open to debate and challenges with no foreseeable solution or answer. Perhaps this was due to his feelings that, while there was plenty of data concerning the nature of “viruses,” the accumulated data was “distinctly lacking in quality,” and that “enough reliable data have not been acquired to establish the nature of the viruses.”
The Nature of Viruses
Thomas M. Rivers
The Rockefeller Institute for Medical Research, New York
Viruses are usually characterized by three negative properties, namely, invisibility by ordinary microscopic methods, failure to be retained by filters impervious to well-known bacteria, and inability to propagate themselves in the absence of susceptible cells. I prefer a positive characterization of the viruses, one emphasizing the intimate relation that exists between them and their host cells. The multiplication of viruses only in the presence of susceptible cells, their regeneration and production of disease in many instances in only one species of host, the marked stimulation and destruction of cells induced by their activity which on the one hand gives rise to tumors, such as Rous’ sarcoma, and on the other to vesicular lesions, as fever blisters, the intracellular pathology frequently evidenced in virus diseases by inclusion bodies, and, finally, the lasting immunity that follows the majority of virus maladies, are essential phenomena that serve to stress the intimate type of parasitism encountered in working with these active agents. Such a characterization of viruses implies much, not only as concerns their biological nature which is still a moot question, but as regards their activities about which something is definitely known.
Data concerning the nature of viruses are sufficiently adequate in quantity but distinctly lacking in quality. According to reports, some of which have come from eminent investigators, most of these active agents have been seen and have been cultivated on lifeless media. If such statements are correct, viruses are autonomous living agents, and further discussion of their biological nature should deal with their place in the scale of living entities and their relation to other forms of life. Reports of work in this field are confusing, however, particularly to the uninitiated, and critical investigators are of the opinion that enough reliable data have not been acquired to establish the nature of the viruses. Inasmuch as this is a subject of fundamental biological importance, I shall review some of the recently accumulated data regarding the size, electrical charge, purification, spontaneous generation, adaptations, elementary bodies, metabolism, immunological phenomena and cultivation of viruses that might be of assistance in the elucidation of the origin and constitution of these peculiar incitants of disease.
One of the only indirect means which early virologists could use to conclude that a “virus” was “present” in a sample was by claiming that the invisible entities passed through filters of a certain size that retained all known bacteria, thus allowing them to guess as to the size of the unseen particles. Rivers noted that a “virus” was generally accepted as “an object less than 0.2 p or 200 ppl in diameter” and that it was not capable of being seen under light microscopy. In other words, “viruses” were too small to be seen and were defined by their absence. He noted that figures regarding the size of “viruses” derived from stained preparations were apt to be inaccurate and misleading. This lines up with his 1927 statement on filtration in his paper Filterable Viruses: A Critical Review, claiming that the methods were “crude and inaccurate.”
Rivers then presented evidence for the size of eight “viruses,” which were contradictory depending upon the researchers cited. He utilized hemoglobin as a comparison and stated that if the figure for hemoglobin is incorrect (which had contradictory estimates as to its size as well), many statements concerning the size of “viruses” were also inaccurate. Rivers was dismayed that certain researchers did not account for the possibility that they might have been estimating the size of particles of degraded cells to which the “viruses” were attached. He noted that other researchers took this into consideration and that they were unable to be assured that they had been successful in obtaining the correct figures for the size of the different “viruses.” Rivers concluded that none of the figures could be accepted without reservations and that the exact size of any “virus” was unknown. The numerous contradictory results stemmed from “inadequate experimentation, careless thinking, prejudice, imperfect experimental methods, and the difficult nature of the problems.”
SIZE. The size of minute particles may be determined in several ways, namely, by direct mensuration provided the objects are capable of resolution under the microscope; by filtration and ultrafiltration if the factors that influence the passage of the particles through pores of graded diameters are known and controlled; by diffusion, and, finally, by centrifugation. All of these methods have been employed in the study of the magnitude of viruses and the results obtained will be discussed.
It is generally accepted that an object less than 0.2 p or 200 ppl in diameter is not capable of resolution under the microscope when ordinary light is used. Furthermore, it is understood that mordants and stains usually increase the magnitude of small particles. Some of the “larger” viruses, e.g., those of fowl-pox (log), smallpox, vaccinia (122, 123), and rabies, are said to be just visible after treatment with certain mordants and dyes. Consequently, one is justified in concluding that most of the viruses have a diameter of less than 200 pp and in an unstained state are not mensurable by means of ordinary light. Moreover, figures regarding their size derived from stained preparations are apt to be inaccurate and misleading. The use of light of short wave lengths makes possible the mensuration of particles smaller than 0.2 p in diameter. So far, however, this method of investigation has yielded no convincing evidence concerning the magnitude of viruses. It appears, therefore, that direct methods of mensuration only indicate that the active agents are considerably smaller than ordinary bacteria.
The sizes of at least eight viruses have been estimated by means of ultrafiltration, diffusion, or centrifugation. The results obtained for these active agents together with figures for the diameter of the hemoglobin molecule for comparison are given below.
Hemoglobin. For a number of years the molecule of hemoglobin was thought to be 30 uu in diameter. Recently, however, figures (34) derived from the results of Svedberg and Nichol’s (33) centrifugation experiments and Northrop and Anson’s (30) diffusion experiments with hemoglobin indicate that its diameter is approximately 5.5 uu. Many estimations regarding the magnitude of viruses have been based on the former figure for the diameter of the hemoglobin molecule, 30 uu. If this figure is incorrect, many statements concerning the size of viruses are also inaccurateMosaic virus. Duggar and Karrer (17) by means of ultrafiltration found the infectious particles of mosaic virus to be of the same order of magnitude as hemoglobin molecules, namely, 30 uu in diameter. Vinson (34), however, says that Duggar’s experiments interpreted in the light of recent work regarding the size of hemoglobin molecules indicates that the diameter of mosaic virus is about 5.5 uu.
Herpetic virus. Zinsser and Tang (38) by means of ultrafiltration estimated the diameter of herpetic virus to be 20-100 uu. Levaditi and Nicolau (27) in the same way found that the virus passed through membranes which retained toxins, hemolysins, complement, and serum globulins. Bedson (13), however, unable to confirm Levaditi and Nicolau’s (27) results, obtained evidence by centrifugation that herpetic virus is probably of sufficient size to be visible.
Foot-and-mouth disease virus. Olitsky and Boez (31), using ultrafiltration, found that the virus of foot-and-mouth disease is 20-100 uu in diameter. Elford by means of his special membranes estimated it to be 8-12 uu.
Poliomyelitic virus. By means of ultrafiltration, Krueger and Schultz (25), in 1929, found that the virus of poliomyelitis possesses a magnitude not greater than 300 uu. In 1931, by the same means, Clifton, Schultz, and Gebhardt (16) obtained results indicating that the diameter of the virus lies below 50 uu.
Fowl plague virus. By means of ultrafiltration Andriewsky (8) secured a figure of 2.5 uu for the diameter of fowl plaguevirus, while Bechhold and Schlesinger (11) by centrifugation found it to be 120-130 uu.
Bacteriophage. According to d’Herelle (22) and Elford (19), both of whom used ultrafiltration, the diameter of the bacteriophage is approximately 30 uu. Kruger and Tamada (26) by means of purified bacteriophage preparations and ultrafiltration found it to be 5 uu, and Hetler and Bronfenbrenner (24) by means of a diffusion method estimated it to be 1.2-22.8 uu.
Rous virus. According to Zinsser and Tang (38), the Rous virus is 20-100 uu; according to Mendelsohn, Clifton and Lewis (29), 50 uu; according to Frankel (20), 10 uu. All these workers obtained their figures by means of ultrafiltration.
Vaccine virus. Levaditi and Nicolau (27) reported that vaccine virus passes through membranes impervious to toxins, hemolysins, complement, and serum globulins. Bland (15), however, from the results of his centrifugation experiments not only concluded that Levaditi and Nicolau’s findings are incorrect but that vaccine virus is probably large enough to be seen. Bechhold and Schlesinger (11) by means of centrifugation estimated that the active agent is 210-230 uu in diameter, while Yaoi and Kasai (37) working with “purified” virus found that it diffused at the rate of fuchsin particles and is, therefore, not capable of being seen.
One cannot consider the results cited above without being amused and dismayed. Many of the workers seemed in no way concerned about the possibility that they might have been estimating not the magnitude of viruses, but the size of particles of degraded cells to which the viruses were attached. Other investigators, cognizant of the difficulties of the problem! attempted to remove the viruses from such carriers. They were unable, however, to be assured that they had been successful and that they had obtained the correct figures for the size of the different viruses.
From the results of indirect methods of mensuration it is safe to conclude that viruses are small and that some of them may be exceedingly minute. If the figure of 210 uu for the diameter of vaccine virus is accurate, there is no reason as far as size is concerned to suppose that the virus is not a living organism. On the other hand, if the figures of 1.2 uu, 5.5 uu, and 8 uu for the bacteriophage, mosaic virus, and foot-and-mouth disease virus, respectively, are correct, it is obvious that these agents cannot be highly organised, because it is impossible that with such a magnitude they can consist of more than one, or at most several, molecules of protein. Unfortunately, none of the figures can be accepted without reservations. At present the exact size of
The numerous discordant results encountered in the literature dealing with the filterability, size, and visibility of viruses are probably due to inadequate experimentation, careless thinking, prejudice, imperfect experimental methods, and the difficult nature of the problems. One of the great needs at present is improvement in methods of microscopy, filtration, and purification of viruses in order that results obtained will approximate the true size of viruses and not the size of particles of other sorts on which the agents are adsorbed. It must be remembered, however, that the determination of the size of one virus will not establish the magnitude of another, because no more uniformity of dimensions should be expected among these agents than is found among bacteria and protozoa. Furthermore, it is not possible to derive proof of the animate or inanimate nature of viruses even from a correct estimation of their diameters, for, within limits as yet undetermined, life and death are not functions of size.
In this next section on electrical charges, take note once again of the range in estimates and contradictory conclusions made by the researchers. Rivers pointed out that “virus” preparations consisted principally of proteins and bits of degraded cells from the host. This meant that the electrical charge results might not be those of the “virus” particles themselves but of the other materials present within the sample. This inability to distinguish the assumed “virus” from the remaining host and foreign constituents present in the sample is the reason why complete purification and isolation of the assumed “viral” particles from the host components, which has never been achieved, is absolutely necessary. Rivers admitted that there were few experiments that were performed with “protein-free” preparations of “viruses,” and that the methods of purification did not convince him that such purified “viruses” had ever been completely separated from their carriers (i.e. host materials). Even with the modern advances in technology, this inability to completely separate “viruses” from host components was noted in a May 2020 article that stated that “to date, a reliable method that can actually guarantee a complete separation does not exist.” Rivers concluded that the lack of purity meant that it was impossible to state definitely what electrical charge is carried by the “viruses.”
ELECTRICAL CHARGE. Most bacteria and proteins under ordinary biological conditions of hydrogen ion concentration carry a negative electrical charge. When the nature of the viruses became a question of interest, attempts were made to ascertain their behavior in an electrical field in order that it might be compared with the action of proteins and bacteria under similar circumstances.
Bacteriophage. Kligler and his co-workers (41) using a so-called “protein-free” bacteriophage found that the active agent was amphoteric in acid and decidedly alkaline solutions and chiefly negatively charged in neutral and mildly alkaline solutions. Krueger and his associates (42) stated that the bacteriophage is negatively charged between the hydrogen ion concentrations of 9.0-3.4, and positively charged at pH 3.35. Todd (48) found that the active agent carried a negative charge between the hydrogen ion concentrations of 3.36-7.6. The results of Natarajan and Hyde’s (43) experiments indicate (1) that bacteriophages for typhoid bacilli and Flexner’s dysentery bacilli are only electronegative between pH 4.9-9.3 and 5.4-9.3 respectively, (2) that small plaque coliphage is electronegative below pH 8.3, but with greater alkalinity moves to both poles, and (3) that large plaque coliphage is electronegative over a range of pH 5.4-6.1, while at a higher alkalinity it wanders to both poles.
Rabic virus. According to Glusman (40) and his associates fixed rabic virus is negatively charged over a range of pH 6.0-9.3.
Vaccine virus. Douglas and Smith (39) found that vaccine virus carried a negative charge between the hydrogen ion concentrations of 5.5-8.4. The experiments of Yaoi and Kasai (49) revealed that between pH 6-7 more virus collected at the positive than at the negative pole, and between pH 8-9 the active agent was demonstrable only at the anode.
Fowl-pox virus. Kligler and his co-workers (41) found fowl-pox virus in “protein-free” preparations to be positively charged on the acid side, amphoteric in neutral solutions, and negatively charged in alkaline solutions. According to Natarajan and Hyde (43), the active agent is amphoteric over a range of pH 6.4-9.3.
Foot-and-mouth disease virus. Olitsky and Bo& (44) believe that the virus of foot-and-mouth disease is positively charged, while Sichert-Modrow (47) is of the opinion that the active agent carries a negative charge over a range of pH 7.0-8.1.
Pcliomyelitic virus. According to Olitsky, Rhoads, and Long (45) poliomyelitic virus wanders to the anode.
Rous virus. Pulcher (46) found that the Rous virus was adsorbed on electropositive and not on electronegative hemoglobins and concluded that the active agent is negatively charged.
Virus of infectious myxomatosis of rabbits. According to Natarajan and Hyde (43), the virus of infectious myxomatosis of rabbits is electronegative over a range of pH 4.9-9.3.
Herpetic virus. Natarajan and Hyde (43) found herpetic virus to be electronegatively charged only between the hydrogen ion concentrations of 7.0-8.9.
From the results of the work cited above it is obvious that most workers have found that under ordinary biological conditions of hydrogen ion concentration certain viruses in an electrical field wander to the anode. Moreover, many investigators have stated that the viruses under these conditions are negatively charged and in this respect are similar to bacteria, cells, and numerous proteins. Others, however, aware of the fact that virus preparations usually consist principally of proteins and bits of degraded cells from the host, realize that the electrical charges determined might not be those of the virus particles themselves but of their carriers, i.e., material on which the virus particles are adsorbed. It is true that a few experiments have been performed with “protein-free” preparations of viruses. But an examination of the methods of purification fails to convince one that such purified viruses had been completely separated from their carriers. Therefore, at present it is impossible to state definitely what electrical charge is carried by the viruses.
This next section is probably my favorite of the entire paper as Rivers sums up the purification problem perfectly. He started off by admitting that “virus-containing” emulsions consisted chiefly of substances unrelated to the “virus.” Thus, he stated that researchers needed to attempt to obtain the “viruses” either in a pure or in a relatively pure state as it was realized that purified “viruses” are essential for the proper study of problems in the field. These problems related to the aforementioned estimation of the size of “viruses” and the determination of the electrical charge, as well as any investigation into the “immunological” responses attributed to “viruses.” He shared a quote by Murphy who, in working to purify the Rous sarcoma “virus” through various manipulative purification processes, stated that it was “hardly conceivable that the active fraction” obtained after these processes could “carry with it through all these manipulations any living organism or virus.” Murphy felt that he was dealing with an enzyme rather than a “virus.” Rivers then backed up his own assertion from five years earlier in 1927 that “No virus had been obtained in an absoutely pure state” by reiterating that it was unlikely that a “virus” had ever been obtained in a state of absolute purity.
PURIFICATION. Inasmuch as virus-containing emulsions consist chiefly of substances unrelated to the active agents themselves, it is natural that workers should attempt to obtain the viruses either in a pure or in a relatively pure state. Moreover, it is being realized that purified viruses are essential for the proper study of problems in this field, such as the estimatlion of the size of viruses, the determination of their electrical charge, and the investigation of immunological responses excited by them. In addition to the fact that purified viruses are of practical value, it is obvious that such preparations will also be of value to investigators interested in the theoretical problem of the nature of viruses. Indeed, Murphy (52, 55) has already concluded from the results of his experiments on the purification of the Rous agent that this disease-incitant is neither a virus nor a living organism. He states, “It is hardly conceivable that the active fraction which I have thus succeeded in obtaining, a substance purified by repeated precipitations, could carry with it through all these manipulations any living organism or virus. To me the enzyme-like nature of the principle seems to have been conclusively established. . . . .” However, most workers do not believe that Murphy is justified in concluding from the results of such experiments that the Rous agent is not a virus, because at least eight other viruses, e.g., the incitants of infectious myxomatosis of rabbits (58), foot-and-mouth disease (64)) bacteriophagy (50, 56, 57), fowl-pox (56), vaccinia (66, 68)) rabies (66), poliomyelitis (65), and mosaic disease (67) have been subjected to manipulations similar to those used by Murphy and have been obtained, still active, in various states of purity.
Most methods of purification of viruses are based on the principles of precipitation by a variety of chemicals and selective adsorption and elution as used extensively in enzyme work. As yet, it is unlikely that a virus has been obtained in a state of absolute purity. Nevertheless, the results already secured are encouraging and should excite further investigations. It may be possible in this way to attain eventually a more accurate concept of the nature of some viruses. For instance, it may be shown that in certain purified virus preparations the number of nitrogen atoms for each infectious unit or particle is insufficient to warrant the supposition that the agents are living, organized structures. Krueger and Tamada (57) have already suggested this viewpoint.
While it is now stated that “viruses” require a host cell and must be cultured in order to be observed and studied, in the past, claims were made that “viruses” could be grown without cells. Rivers stated that these claims of successful cultivation on lifeless media were not uncommon, and he noted a few cases:
- Frosch and Dahmen stated that they were able to cultivate the “virus” of foot-and-mouth disease on ordinary media.
- Olitsky reported the cultivation of mosaic “virus” in a cell-free medium.
- Eagles and McClean reported that vaccine “virus” is capable of regeneration in a cell-free medium.
Rivers ultimately decided that none of these were true examples of “viruses” being grown in cell-free media, and thus, it was chalked up to contradictory evidence that was brushed aside in favor of the prevailing belief that “viruses” are invisible and incapable of regeneration in the absence of living susceptible host cells. Rivers did note that such a state of affairs would prevent a complete definition of the nature of “viruses.” However, he believed that it was not absolutely essential to see and to cultivate the “viruses” on simple media.
CULTIVATING. In the literature of twenty years ago it is not uncommon to encounter reports in which it was claimed that viruses had been successfully cultivated on lifeless media. These reports have not been confirmed and at present such claims are rarely made. A few, however, have been made in recent years. Frosch and Dahmen (78) stated that they were able to cultivate the virus of foot-and-mouth disease on ordinary media. But the German, English, and American Foot-and-Mouth Disease Commissions were unable to confirm their work. Olitsky (91) reported the cultivation of mosaic virus in a cell-free medium. Nevertheless, upon repeating his work he (92) has been forced to conclude that true multiplication of the virus was not obtained. Recently, Eagles and McClean (75, 76) reported that vaccine virus is capable of regeneration in a cell-free medium. A careful examination of their papers, however, leaves one in doubt as to whether some of their media were cell-free, and as to whether multiplication of the virus occurred in the nutrient materials that undoubtedly contained no cells. In my laboratory (86, 90, 93) during the last four years, Haagen, Muckenfuss, Li, and I have made numerous attempts to cultivate vaccine virus in cell-free media, many of which were similar to if not identical with those employed by Eagles and McClean. None of our efforts was successful. On the other hand, the cultivation of vaccine virus in the presence of cells surviving in vitro has been more consistently successful in our hands and in Maitland’s (88) than it has been in Eagles and McClean’s.
Although the cultivation of viruses in lifeless media has not been accomplished, it is generally conceded that these agents are capable of pullulation in the presence of susceptible cells either surviving or growing in vitro. The viruses of Rous sarcoma (72), Virus III infection of rabbits (69), herpes febrilis (70), fowl-pox (77), vaccinia (79, 80, 86, 88)) rabies (94), foot-and-mouth disease (83,84,85? 89), vesicular stomatitis (73), infectious myxomatosis of rabbits (71, Sl), fowl plague (82), and probably the agents causing common colds (74) and poliomyelitis (87), have been cultivated in the presence of tissues surviving in vitro.
Moreover, the characteristic of species specificity possessed by many viruses is frequently reflected in their in vitro cultivation. For instance, fowl-pox virus (77), innocuous for mice and rats, does not regenerate in cultures of their tissues. Foot-and-mouth disease does not attack chickens and the virus (89) does not grow in cultures consisting of minced chick embryo and plasma. In addition to a species specificity, some viruses exhibit in cultivation experiments a predilection for certain kinds of cells. Fowl plague virus (82) multiplies in the presence of chick embryo skin and brain, but does not regenerate in pure cultures of fibroblasts. Foot-and-mouth disease virus (85) increases in amount when the culture medium contains minced guinea-pig embryo, but does not grow when fibroblasts or bits of heart muscle alone are present. Thus it appears that many viruses are capable of multiplication in tissue cultures and frequently retain under such conditions their species and cellular specificity. Nevertheless, it will be interesting to observe the results of further attempts to circumvent this species and cellular specificity of viruses by in vitro methods of cultivation.
A crucial experiment, if there be one, to decide the question of the autonomy of the viruses is their undisputed cultivation on lifeless media. It may be impossible, however, to accomplish such an experiment with all of the viruses, because some of them may be obligate parasites, as is the malarial organism. Thus in the quest for proof of the nature of viruses, we may find that many of them are invisible and incapable of regeneration in the absence of living susceptible host cells. Such a state of affairs will prevent, for a time at least, a complete definition of the nature of these peculiar incitants of disease. Nevertheless, we should obtain all the facts and make the most of them in the study of biological phenomena and in the better understanding and control of disease. For this purpose it is not absolutely essential to see and to cultivate the viruses on simple media any more than it is imperative to see and to know what electricity is in order to study the phenomena produced by it and to control its activity for our daily needs.
Regarding whether “viruses” are alive or not due to having their own metabolism, Rivers stated that the evidence was that they did not have any such metabolic capabilities. However, he felt that conclusions could not be drawn that “viruses” do not have a metabolism and that they are inanimate because the methods used may not have been adequate.
Adaptation of the “virus” to different hosts was used by researchers in order to state whether or not “viruses” were alive. This essentially meant drawing conclusions from using different materials and different methods in different animals while generating different results. One set of researchers viewed the contradicting outcomes as the result of a “living virus” while another set of researchers saw it as a result of the hosts response. Neither seemed to recognize the fact that it was the different experimental procedures generating different responses and results rather than the act of any “virus” adapting.
METABOLISM. Much of the discussion conc.erning the nature of viruses has centered around the question as to whether they are animate or inanimate. In this relation, one would like to know what the evidence is regarding independent metabolic activities of these active agents. Technical difficulties have hindered this type of experimentation with viruses. Nevertheless, a few investigations (95-99) have been made, the result,s of which were negative. One must not conclude from such negative results, however, that viruses do not possess an independent metabolism and are, therefore, inanimate substances, because the methods used for the detection of the metabolic activities may not have been sufficiently delicate.
ADAPTATION. Certain viruses inoculated into new hosts apparently undergo changes in some of their characteristics. Smallpox virus (100, 101) passed through monkeys to rabbits and calves and then back to man is no longer smallpox virus but vaccine virus, a.nd the disease, vaccinia, caused by it is not contagious as is smallpox. The incitant of yellow fever (106, 107) passed through a large number of mice by means of intracerebral inoculations loses much of its pathogenicity for monkeys when inoculated intravenously or intraperitoneally, but gains the power of producing a transmissible encephalitis in monkeys receiving the inoculum in the brain. Such phenomena are spoken of as adaptations of viruses to new hosts, and, inasmuch as adaptation is considered a characteristic of living rather than lifeless material, they have been cited by some investigators (103) as proof of the animate nature of the viruses. On the other hand, workers, who believe that viruses are products of cellular perversion, state that the changes observed in the characteristics of the active agents when they are inoculated into alien hosts are to be expected, inasmuch as mouse, rabbit, monkey, and human cells, because of intrinsic differences, may not always manufacture identical substances as the result of similar stimuli. Therefore, they contend that the changes and adaptations are not accomplished by the agents themselves but by their hosts and, consequently, are not admissible as proof of the living nature of the viruses.
As ”viruses” were incapable of being observed and studied directly, various forms of indirect evidence were utilized in order to infer the presence of these entities. One of the earliest ways to do so was by claiming that a phenomenon known as inclusion bodies was a sign that a “virus” was present. These “bodies” are aggregates of proteins seen in various tissues under microscopy that were taken as an indicator by the researchers that they were dealing with a “virus.” However, it is well-known that inclusion bodies are not specific to “viral” cases and can be found in those without a “viral” disease. They are also not found in all cases of a particular disease, can be found in those without the disease, and are even found in uninoculated cell cultures, as seen with RSV. A 1941 paper by Alfred M. Lucas stated that the “existence of an object which appears to be an inclusion body is not proof of the presence of a virus but merely an indication that a virus should be considered if no bacterial agent can be found.” What this means is that inclusion bodies are nothing more than non-specific indirect evidence used to infer an assumed “virus” if other “causes” are ruled out. This means that finding inclusion bodies is essentially meaningless as a specific sign for the presence of any “virus.” Rivers appeared to understand this as well. After presenting various contradictory interpretations and presentations of inclusion bodies by different researchers, he noted that “inclusions may arise in a number of ways and that they may or may not contain virus.” He felt that making conclusions about what these “peculiar structures” represented was “hazardous at present.”
INCLUSIONS. Within the nucleus and cytoplasm of cells injured by viruses, certain peculiar structures, inclusion bodies, are frequently observed. Although many of these bodies are of importance in diagnostic and experimental work, numerous opinions exist concerning their nature. Lipschtitz believes that the inclusions in many diseases (119) consist of compact masses of virus particles, yet he is of the opinion that such structures in measles (120) are nothing more than altered central bodies. Goodpasture (113) thinks that Negri bodies in rabies are composed of degenerated mitochondria and neurofibrils, while Levaditi (118) and Manouelian (121) consider them protozoa and designate them, respectively, Glugea lyssae and Encephalitoxoon rabiei. Goodpasture and his associates (124, 125) have demonstrated that the incitant of fowl-pox is intimately associated with the Bollinger bodies which are made up of a lipoid capsule within which numerous small coccoid bodies are embedded in a protein matrix. On the other hand, Glaser (112) has presented evidence that the polyhedral bodies, the characteristic inclusions in wilt diseases of caterpillars, consist of non-infectious crystalline protein. Thus, it appears that inclusions may arise in a number of ways and that they may or may not contain virus. Consequently, generalizations regarding these peculiar structures are hazardous at present.
The small coccoid bodies found in fowl-pox by Borrel (109) and in vaccinia by Paschen (122,123) appear to be extremely minute organisms. In fact, one is justified in asking why these bodies are not convincing evidence of the organismal nature of certain viruses. The first reason is that one cannot by morphological and tinctorial data alone determine whether autonomous life exists in such small objects. Another reason is the fact that Goodpasture, while holding the belief that the small coccoid bodies in fowl-pox (124, 125) represent the virus, stated that similar structures, seen in rabic brains (113) and considered of etiological importance by Babes (108) and Koch (114-116), are probably degenerated mitochondria. Moreover, Borrel (110) has described similar bodies in other virus diseases the etiological agents of which have been shown by ultrafiltration to be incapable of resolution by microscopic methods. Furthermore, Craciun and Oppenheimer (111)) who cultivated the small bodies of vaccinia and showed that they are closely associated with the virus, made the following statement, “We have from these studies no morphological proof of an increase in the number of granules, since they cannot readily be distinguished from other granules normally seen in tissue cultures.” Finally, mitochondria in some respects resemble bacteria. They may decrease or increase numerically within cells, and their size and shape may be altered by appropriate stimuli. At times, they actually divide. Nevertheless, mitochondria are not considered autonomous living agents. Consequently, so far as I am aware, there is no convincing evidence-the specific agglutination of virus elementary bodies (184) by antiviral sera will be discussed later-to invalidate the conception that cells under the stimulus of viruses may react by the formation of numerous small coccoid bodies uniform in size and intimately associated with the stimulating agents. One would not consider such bodies microorganisms or hold that they consist of virus alone. Therefore, in spite of definite proof that viruses are present in certain types of inclusions, doubt still exists regarding the organismal nature of the small coccoid bodies found within them.
Other features observed in pathological processes induced by viruses, e.g., hyperplasia and necrosis, are fully as important as are the inclusion bodies. The excessive stimulation of cells seen in some virus diseases, e.g., fowl-pox and warts, leads one by analogy to think of mdignant neoplasms. Undoubtedly a number of fowl tumors are caused by agents separable from cells, and, although there is no proof that mammalian tumors arise in this way, the possibility is worthy of consideration and offers an attractive field for work. The fact, however, that some tumors are produced by filterable agents is by no means conclusive evidence that all neoplasms (217) arise through the activity of such incitants.
In this next section, Rivers admitted that there was an increasing chorus of researchers who believed that “viruses” were nothing more than “merely filterable, invisible, and noncultivable elements of ordinary bacteria.” He presented many scenarios, such as:
- The bacteriophage is a form in the life cycle of lysogenic bacteria.
- The “viruses” of yellow fever and hog cholera are invisible forms of Leptospira icteroides and B. suipestifer respectively.
- The etiological agent of scarlet fever is a filterable form of hemolytic streptococci.
- The incitants of poliomyelitis, epidemic encephalitis, fox encephalitis, common colds, measles, and influenza represent certain stages in the life cycle of green streptococci.
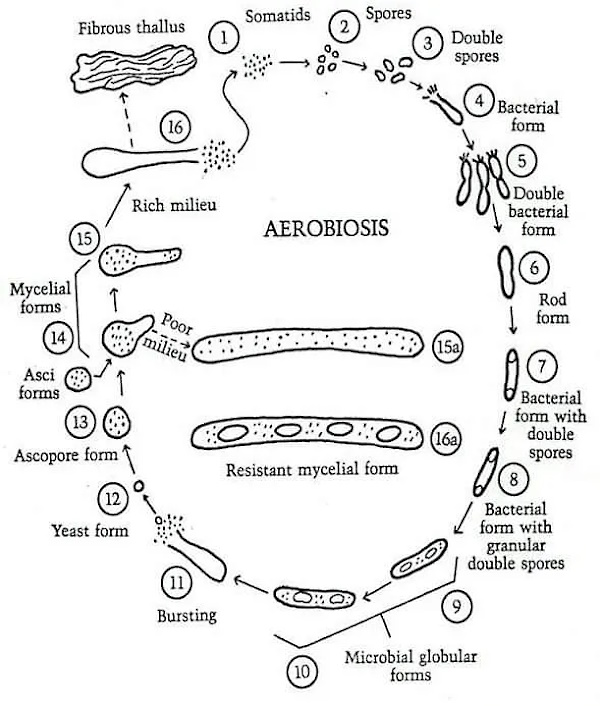
Apparently, Rivers was unfamiliar with the fact that this bacterial life cycle process, known as pleomorphism, was observed by many researchers such as Antoine Bechamp, Günther Enderlein, Royal Raymond Rife, and later by many others with the use of dark field microscopy. That bacteria are pleomorphic entities, i.e. having the ability to assume different forms, is an established fact.
VIRUSES AS FILTERABLE FORMS OF BACTERIA. For a long time a few investigators have held that certain virus diseases are induced by ordinary bacteria. Now that attention is being focused on filterable forms of bacteria, workers in increasing numbers (128, 131, 132, 134, 135) are adopting the belief that viruses are merely filterable, invisible, and noncultivable elements of ordinary bacteria. It has been claimed, and evidence of a kind has been offered to substantiate the assertions, that the bacteriophage (165, 166) is a form in the life cycle of lysogenic bacteria, that the viruses of yellow fever (131, 134, 135) and hog cholera (134, 135) are invisible forms of Leptospira icteroides and B. suipestifer respectively, that the etiological agent of scarlet fever (134, 135) is a filterable form of hemolytic streptococci, and that the incitants of poliomyelitis, epidemic encephalitis, fox encephalitis, common colds, measles, and influenza represent certain stages in the life cycle of green streptococci (131). Without going into details of the available knowledge of bacterial life cycles and their invisible and noncultivable forms, one can say that proof of many of the claims regarding them is lacking. In fact, if certain reports are correct, some of the filterable forms of bacteria are much smaller than are many of the viruses. Kendall (131) recently stated that “egg white, filtered through Berkefeld W filters (after dilution with sterile physiological saline solution) is rarely sterile.” Such a statement raises embarrassing questions for workers in the virus field because many viruses will not pass through W filters. Since the existence of bacterial life cycles is doubtful, it seems unwarrantable to offer the presumptive filterable forms of them as evidence upon another unsolved problem, the nature of the viruses.
The thing to notice in this next section on physical and chemical agents is, once again, the often contradictory nature of the evidence presented by different researchers. One researcher would find a certain chemical that had an effect on the “virus,” while another researcher would state otherwise. Some viewed that chemical tests proved “viruses” were protozoa. Others felt that their tests proved the “virus” was an enzyme. Sanderson showed that bacteriophages were not killed by successive freezing and thawings and believed that they were unliving. However, Rivers showed that bacteriophages can be killed by repeated freezing and thawing, thus contradicting Sanderson’s interpretation. Ultimately, Rivers concluded that, regardless of the number of tests with chemical and physical agents that had been devised as criteria for the presence of life or to define the nature of “viruses,” not a single one of them was found to be satisfactory.
EFFECT OF PHYSICAL AND CHEMICAL AGENTS ON VIRUSES. Many years ago it was discovered that bile and saponin are injurious to protozoa but with a few exceptions are innocuous for bacteria. Consequently, when the question of the nature of viruses began to attract attention, tests were made to determine what effect bile and saponin have on these incitants of disease. Many viruses, e.g., rabic virus (141, 144), were found to be inactivated and because of this fact certain workers concluded that they are protozoa. Sufficient exceptions, however, have been encountered to invalidate the test as a means either of separating bacteria from protozoa or of defining the nature of viruses. The agent causing Rous’ sarcoma (140) is more resistant to ultraviolet light than are bacteria, and Murphy (220) considers this fact as evidence in favor of his hypothesis of the enzyme-like nature of the virus. On the other hand, bacteriophage (139), the living nature of which many doubt, is just as sensitive to ultraviolet light as are bacteria. Sanderson (153), using a temperature of -78°C., found no diminution in the titer of two strains of bacteriophage subjected to 20 successive freezings and thawings. Since bacteria and cells are killed by repeated freezing and thawing, he concluded that bacteriophage must be something other than a living organism. Rivers (151) showed, however, that colon bacilli, Virus III, vaccine virus, herpetic virus, bacteriophage, complement, and trypsin are all either killed or inactivated by repeated freezing (-185°C.) and thawing and that, as might be expected, some of the agents are more resistant than are others. Hence it is obvious that destruction or inactivation of an active agent by repeated freezing and thawing is not evidence that it possesses life. The observations on heat, desiccation, oxidation, and the effect of dyes have likewise yielded no convincing evidence concerning the nature of viruses. Thus it appears that a number of tests with chemical and physical agents have been devised as criteria for the presence of life or to define the nature of viruses, but no one of them has been found satisfactory.
The spontaneous generation of “viruses” by the host is a concept that defeats the idea that these entities are exogenous outside invaders. If something like a bacteriophage can be produced by a normal bacterium without any external phage present, it shows that these entities arise from a process initiated from within the organism. Rivers noted that Hadley and his co-workers stated that it was possible to obtain bacteriophage from normal bacterial cultures by means of enforced dissociation. Thus, no external source of phage was necessary. Nobel Prize-winning immunologist Jules Bordet was able to do the same, as did other researchers. Rivers presented a few scenarios where “viral” diseases could be induced by injecting toxic substances such as tar and arsenic into chickens, as well as an instance where a tumor-producing extract could be obtained from healthy chickens. While Rivers thought that the interpretation of the evidence was potentially fundamental to biology, he excused it as being due to contamination by the researchers working in labs with similar materials as well as the possibility that “latent viruses” were hiding within the healthy hosts.
SPONTANEOUS GENERATION OF VIRUSES. The origin as well as the nature of viruses constitutes a question of interest. The intimate relation between these active agents and their host cells has induced more than one investigator to view the host cell as the source or origin of viruses. Indeed, reports of experimental work have appeared leading to claims that normal cells have been induced to manufacture certain viruses. According to Carrel (156, 157), minced chick embryo mixed with tar, indol, or arsenic and injected into normal chickens in a small percentage of instances gives rise to tumors resembling Rous’ sarcoma no. 1 and transmissible by cell-free filtrates. Fischer (163) by treating cultures of normal cells with arsenic obtained on one occasion a filterable agent capable of causing tumors. Carrel was unable to confirm Fischer’s work. Murphy (52, 167), by means of a method the details of which have not been described, reported that he was able to extract a filterable tumor-producing agent from the gonads of normal-appearing Plymouth Rock roosters. Recently, Hadley and his co-workers (166) stated that it is possible to obtain bacteriophage from normal bacterial cultures by means of enforced dissociation. Although no worker in this field has claimed to have generated living organisms from inanimate matter, it appears that a few believe that they have by certain manipulations induced cells to yield substances which possess some of the attributes of life, notably that of increasing without limit.
The observations described above are suggestive, and, if confirmed and found to warrant the interpretation given them by Carrel, Murphy, Fischer, and Hadley, will prove to be of fundamental biological importance. Unfortunately, however, all of the experiments yielding the observations were actively referred to were conducted in laboratories where workers engaged in the study of agents similar to those supposedly brought into existence. In such laboratories and with such materials it is always difficult for one to rule out the possibility of contaminating normal animals, tissues, bacteria, emulsions, and filtrates. This fact has long been appreciated by workers in vaccine virus laboratories and it delayed the acceptance of the experimental transformation of smallpox virus into vaccine virus. Therefore, experiments of the nature described should never be conducted in rooms used for the study of agents similar to those for which a search is being made. The workers who believe that they have induced viruses to come into existence have not excluded the possibility of the preexistence of latent viruses or of small amounts of virus in the supposedly normal embryos, gonads, chickens, and bacterial cultures utilized in the experiments. This possibility is emphasized by Flexner’s (164) work on poliomyelitis, for he was able to demonstrate the presence of virus in the nasal washings from normal contacts. The possibility outlined is further emphasized by Andrewes and Miller’s (155) experience with Virus III in rabbits, by Cole and Kuttner’s (158) work with the salivary-gland virus in guinea pigs, and by the work upon virus carriers in general among animals, plants (168) and bacteria.
Rivers next discussed “immunity” in relation to establishing the nature of “viruses.” It is important to note that, regarding antibodies and “immunity,” researchers are utilizing one hypothetical entity in order to define another. While Rivers spoke as if the antibody and antigen concepts are established facts, he remarked that if the concept of the nature of antigens is correct, “viruses” are proteins or are closely linked to proteins. Thus, the interpretation of the nature of the “virus” rests upon the correctness of the nature of the antigen concept. He felt that the rise of these (hypothetical) antibodies that differed between host cell and antigen adduced (led one to believe) the exogenous rather than the endogenous origin of the “viruses.” Regardless, Rivers admitted that the mode of action of neutralizing antibodies was not clearly understood, and when speaking of antibodies causing flocculation (clumping together), he shared that various researchers noted the “immunological” phenomena in “virus” maladies are comparable to those induced by toxins. While Rivers felt that “immunological” observations were important, he admitted that this method of approach had not brought about a definite solution to the problem of the nature of “viruses.”
IMMUNITY. Most virus diseases lead to a marked and lasting immunity in recovered hosts. Not only are the but in their sera antibodies capable of hosts refractory to reinfection neutralizing the viruses are demonstrable. What bearing have these facts upon the nature of viruses? In the first place, it is certain that viruses are highly antigenic. Furthermore, if our concept of the nature of antigens is correct, the viruses are proteins or are closely linked to proteins. Moreover, the agents are not only antigenic, but they give rise to antibodies different from those excited by proteins of the host cells. This is true even of the bacteriophage (188). These facts have been adduced as evidence of the exogenous rather than the endogenous origin of the viruses. Thus, the antigenic nature of viruses appears to be prejudicial to the idea that they are products of cellular activity. The notion, however, that a lifeless agent may be injurious to the cell creating it and that it may induce immunological responses independent of those excited by the cell, loses some of its fantastic qualities when one considers the well-known facts that lens protein is not species specific but organ specific and that sympathetic uveitis in the uninjured eye is caused not by microorganisms but by the reaction of the body to substances derived from injured cells of the other uveal tract.
In addition to the neutralizing antibodies, whose mode of action is not clearly understood, complement-fixing antibodies and antibodies causing flocculation in virus emulsions have been described. Schultz and his associates (191-195) contend that the latter types of antibodies are not excited by viruses and that the immunological phenomena in virus maladies are comparable to those induced by toxins. In spite of their contentions, sufficient evidence has been adduced by different workers to make it more than likely that certain virus diseases lead to the production (176, 177, 180, 199) of the antibodies mentioned. Furthermore, Ledingham (184) has recently demonstrated that Borrel bodies in fowl-pox and Paschen bodies in vaccinia are specifically agglutinated by antifowl-pox and antivaccinal sera respectively. The results of these experiments indicate to Ledingham that the elementary bodies are living organisms and represent the virus. There is no reason to doubt that specific agglutinations of the bodies occurred in the manner described by Ledingham, and one cannot deny that such a phenomenon is presumptive evidence of the organismal nature of the bodies. Yet one dare not say categorically that his experiments are unequivocal evidence that the elementary bodies represent virus alone, because it has been shown by Jones (182, 183) that collodion particles treated with a variety of proteins and then thoroughly washed are specifically agglutinated by the proper antisera. Thus, the Borrel and Paschen bodies without being organisms yet having virus adsorbed on them might nevertheless be specifically agglutinated by appropriate antiviral sera.
Gye (181) states that Rous virus repeatedly injected into alien hosts excites two groups of antibodies, one of which acts on the virus itself, while the other operates on the “specific factor” derived from the host cell. According to him, either set of antibodies inactivates the virus. This fact is offered by him as further evidence of the dual nature of the causative agent of fowl tumors. Murphy (189) and Sittenfield (196-198) have reported the presence in Rous sarcoma of a substance that inhibits the action of the etiological agent, and the first mentioned worker is of the opinion that the “inhibitor” differs from ordinary virus antibodies. The presence of this “inhibitor” together with other phenomena has induced Murphy (220) to believe that immunity to the Rous agent is unlike that observed in virus maladies and lends evidence to his view that the Rous agent is not a virus. Inhibiting substances, however, have been obtained from tissues infected with viruses, for example, a substance restraining the action of rabic virus has been demonstrated by Marie (186) in the brains of rabid animals. Furthermore, Andrewes’ (172, 173) work appears to indicate that the immune responses excited by the filterable agents of fowl tumors may not be unique and may possess much in common with those encountered in other virus diseases.
From what has been said, it is obvious that immunological phenomena are playing an important role in discussions concerning the nature of viruses. As yet this method of approach has not brought us to a definite solution of the problem.
Rivers finished up his review on the nature of “viruses” by presenting the various differing interpretations on the concept of the “virus.” These invisible entities were regarded as either:
- Living contagious fluids
- Oxidizing enzymes
- Protozoan parasites
- Inanimate chemical substances
- Minute living organisms (related to bacteria)
Rivers noted that depending on the researchers, the rabies “virus” was either an enzyme, a parasite, a protozoon, or an unknown living organism. He stated that researchers were divided over whether bacteriophages were an inanimate agent or a living organism. The fowl-pox “virus” was thought of as either a protozoan parasite, a nucleoprotein poison manufactured by “infected” cells, or a minute coccoid organism capable of regeneration in parasitized cells. The agent associated with Rous sarcoma was either animate, a living organism mixed with an inanimate substance, an enzyme-like substance, or a transmissible mutagen.
Rivers highlighted these numerous competing concepts in order to show how radically different the ideas concerning the nature of “viruses” are from one another. He then proceeded to explain the main conceptions of “viruses,” with the first two scenarios explaining how a stimulus induces a normal cell to create a substance X, which may either remain free or become closely bound to a part of the cell. In Rivers’ third example, which he considered the most popular, X is a minute living organism that enters cells, multiplies, and produces disease. Thus, there is a distinct difference where X is considered an inanimate substance that results from cellular perversion in the first two scenarios, while X is viewed as an autonomous organism in the last scenario. Regardless of the scenarios that Rivers provided attempting to explain “viral” formation, he admitted that there was no unequivocal evidence of the validity of any of these concepts.
CONCEPTS OF THE NATURE OF VIRUSES. A review of the data by means of which one arrives at a concept of the nature of viruses has been presented. Now it will be interesting to see what notions certain workers have concerning some of them.
Beijerinck (202) considers the virus of mosaic disease to be a living contagious fluid; Woods (228), an oxidizing enzyme; Goldstein (212), a protozoan parasite; Vinson (67), an inanimate chemical substance. Most workers, however, believe that it is a minute living organism.
Hijgyes (216) is of the opinion that the incitant of rabies is an enzyme or “alternatively, that the tissues themselves might spontaneously become virulent as the result of changes in their chemical composition.” At one time Remlinger said, “The rabies virus, which is at once filterable, diffusible and capable of reproducing the disease from case to case, appears to occupy a place midway between the microbes and the diastases.” Recently, however, he (221) has published an article on the evolution of the parasite of rabies. Levaditi (118) and others (121, 225) have presented evidence in favor of the idea that the causal agent is a protozoon. The majority of investigators hold the concept that the incitant is a living organism whose nature is not definitely known.
Numerous workers believe that the bacteriophage is an inanimate agent, while others are convinced that it is a living organism. Ideas, however, concerning the nature of the inanimate transmissible substance or the animate organism vary. For details of the different concepts one is referred to papers by Twort (226, 227), d’Herelle (103), Bordet (203), Bronfenbrenner (95), Burnet (206), and Hadley (165, 166).
The incitant of fowl-pox has been described by certain investigators as a protozoan parasite. Sanfelice (222, 223) suggested that it is a nucleoproteid poison manufactured by infected cells. Borrel (log), Goodpasture (124, 125), and Ledingham (184) hold that it is a minute coccoid organism capable of regeneration in parasitized cells.
Rous and others are prepared to entertain the idea that the causal agent of Chicken Tumor No. I is animate. Gye (215) believes that it consists of two factors, one of which is a living exogenous organism, the other an inanimate specific factor derived from infected cells. Murphy (52, 55), at one time, spoke of the Rous agent as an enzyme-like substance. Recently, however, he (220) has compared it to filterable substances capable of transforming melitensis (204,205) into paramelitensis organisms and of converting one type specific pneumococcus (201) into another type specific form. In regard to the matter he says (22O), “Thus we have a group of agents, products of specialized cells capable of conferring the peculiar type quality to undifferentiated cells of the same species which, in turn, may produce the active factor and transmit this to their descendants.” For this type of agent he proposes the name transmissible mutagens.
Sufficient ideas concerning the nature of viruses have been cited to illustrate how radically some differ from others. Many of them, particularly the ones dealing with the origin and reproduction of inanimate substances that behave in a manner similar to that of living organisms, lack precision. In a general way, however, the different concepts can be arranged in groups and it seems advisable to state and to portray diagrammatically several of the popular ones.
According to one conception, certain stimuli produce changes within cells that are inherited by daughter cells. Once the mutations occur, cells of the new type continue to be formed though the stimuli disappear. No agents separable from the cells are demonstrable, and immunological phenomena in this type of disease differ from those observed in virus maladies. Ordinarily this idea of the causation of disease and the concepts concerning the nature of filterable viruses are not grouped together. Yet in some respects they are not dissimilar and many hold the view that malignant neoplasms arise in some such way. See figure 1.
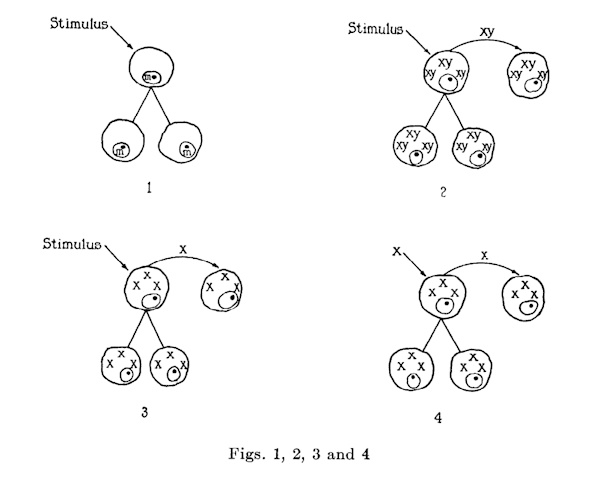
Another notion is that appropriate stimuli induce normal cells to make a substance x which is closely bound to parts y of the cells. Thus an xy complex is formed. This complex, separable from the cells, yet capable of inciting its own production by them, either passes directly into daughter cells, or, having become extracellular, enters another set of normal cells. The xy complex is antigenic, and cells freed from it presumably become normal again. See figure 2.
Still another idea is that certain stimuli incite normal cells to produce a substance x which is not closely bound to parts of the cells, X, separable from cells, yet capable of impelling its formation by them, either passes directly into daughter cells, or, having become extracellular, enters a new group of normal cells. X is antigenic and cells freed from it presumably become normal again. See figure 3.
Finally there is the concept most generally held that x is not a product of the perverted activity of cells but is a minute living organism. X enters cells, multiplies, produces disease, is separable from cells: and is antigenic. Cells freed from it presumably become normal again. At times, x is absorbed by particles y of host cells and evidences of an xy complex are obtained. See figure 4.
For practical purposes it makes little difference which one of the last three concepts is accepted. Theoretically, however, x of the second and third conceptions is quite different from x of the fourth. In the second and third, x, a product of cellular perversion, is an inanimate agent, while in the fourth it is an autonomous organism. No unequivocal evidence of the validity of any of the concepts has been adduced.
Rivers concluded by acknowledging the confused state of the evidence concerning “viruses,” noting that this confusion had made it exceedingly difficult to define their nature. He felt that the easiest way out of their dilemma would be to accept “viruses” as minute organisms. However, Rivers warned of quickly accepting presumptive evidence as “viruses” may be either minute organisms, forms of life unfamiliar to us, inanimate transmissible incitants of disease, or all of the above.
Conclusion
The confused state of our knowledge of the viruses at the present time makes it exceedingly difficult to define the nature of these active agents. The easiest way out of the dilemma, however, would be the acceptance of the presumptive evidence that viruses are minute organisms. Yet the easiest way and the one that best fits the experiences of the day may not be the right one. Furthermore, excessive skepticism and the habit of too readily accepting presumptive evidence are equally productive of sterility. Unless viruses represent a form of life unknown to us, proof of their living nature would not be a striking discovery. If, however, some of them are not animate, absolute proof of such a fact would be of fundamental biological importance. Therefore, care should be exercised that immoderate skepticism on the one hand, and the mental satisfaction secured by accepting presumptive evidence on the other, do not dull our efforts to obtain a better understanding of the viruses, some of which may be minute organisms, while others may represent forms of life unfamiliar to us, while still others may be inanimate transmissible incitants of disease. In any event, we are face to face with the “infinitely small in biology,” and, if there be a sharp demarcation between life and death, then scientists, investigating the nature of viruses, are working near the line that separates infinitely small living organisms from inanimate active agents.
https://www.nature.com/articles/145853d0
From these two presented articles from two different points in time in the history of virology (Rivers in 1932 and Summers in 2014), it should be clear why it is difficult for virologists to define the nature of the “virus.” Researchers needed to invent, and then continually reinvent, the nature of the “virus” as the foundation that virology is built upon is conceptually weak. It is full of contradictions that have cracked the very infrastructure that was put in place. There were never any submicroscopic entities that were being studied by the various researchers over the last century. As there were no “viruses” to study and characterize, there was no agreement at all amongst the various researchers as to the nature of the invisible concept crafted inside of their minds. They had tricked themselves, through shoddy indirect pseudoscientific evidence, into believing that they were studying something real based upon lab-created effects without an identifiable cause. This is why the “virus” has been continually defined for what it isn’t, rather than for what it supposedly is. The magical “virus” skirts the line between life and death, microbe and molecule, enzyme and ferment. It is unlike anything else seen in nature, and for this very reason, its nature remains mysterious and incomplete. This should be the very first clue that there is nothing scientific about the “virus,” as science only deals with the natural world and its phenomena, not the supernatural. However, within the supernatural realm is where the “virus” concept will remain, ready and waiting to be reinvented upon the arrival of the latest technology for the next best indirect measurement. This will be utilized to continue fooling the researchers, as well as the public that blindly trusts in them to know better, that these fictional entities exist in nature, when, as Thomas Rivers kindly pointed out, “viruses” have never once been observed there. Thus, the nature of the “virus” will continue to remain merely an invention of the imagination of the most ardent admirers of these invisible boogeymen—the virologists.
This article originally appeared on ViroLIEgy’s Antiviral Substack.
Connect with Mike Stone website | substack
Cover image credit: flutie8211


Truth Comes to Light highlights writers and video creators who ask the difficult questions while sharing their unique insights and visions.
Everything posted on this site is done in the spirit of conversation. Please do your own research and trust yourself when reading and giving consideration to anything that appears here or anywhere else.








